| Dimetrodon Fossil range: Asselian to Early Kungurian (Permian) | |
|---|---|
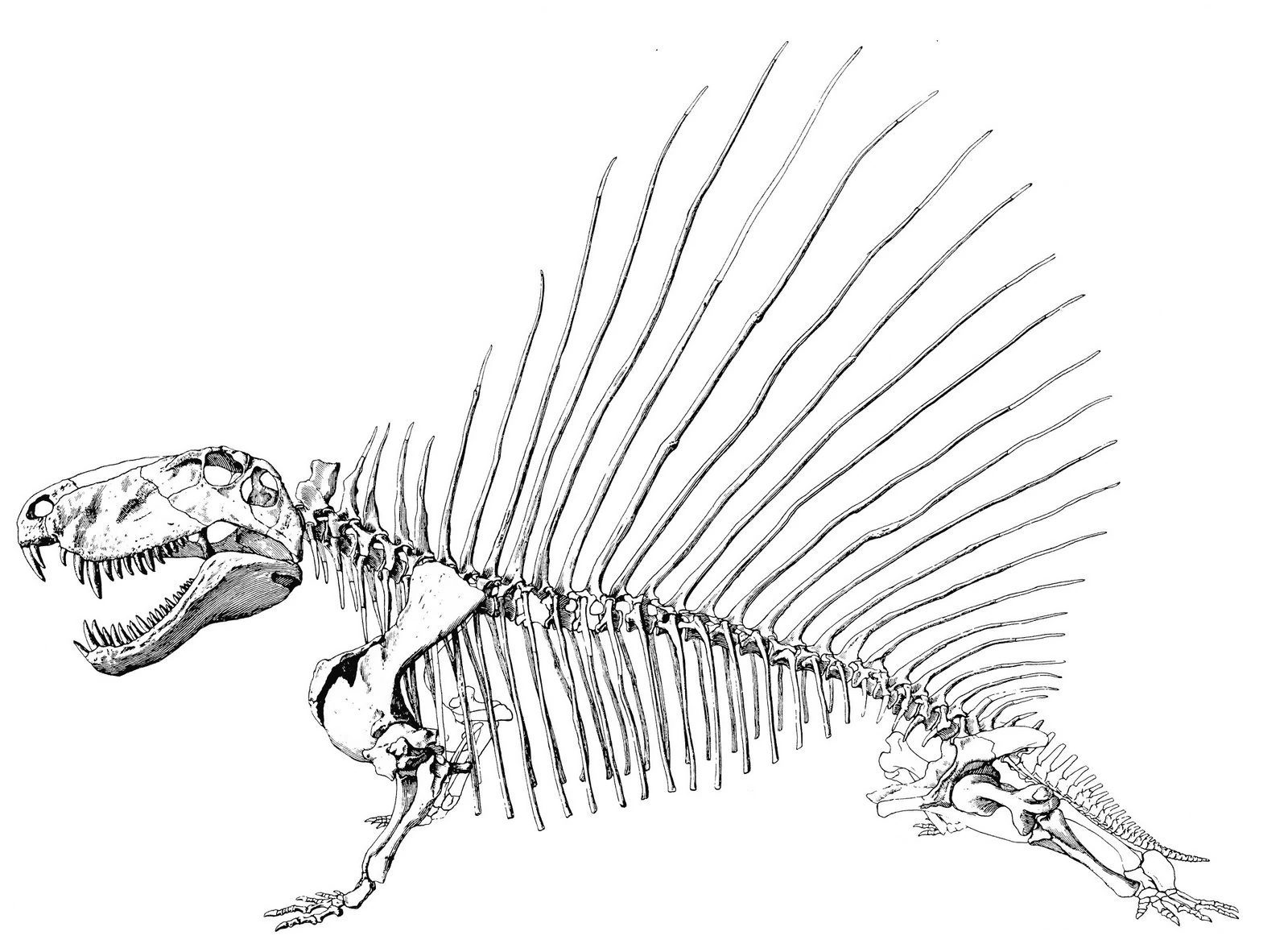 Skeletal diagram of Dimetrodon by paleontologist Robert T. Bakker | |
| Scientific classification
| |
Dimetrodon (meaning "two-measures tooth", in reference to the fact that unlike reptiles, the teeth differ in size.) is an extinct genus of terrestrial predatory synapsid ('mammal-like reptile') genus of pelycosaur that flourished during the Permian Period, living between 280–265 million years ago. Dimetrodon was the largest and most specialized pelycosaur, and remained the dominant carnivore of its environment. It was superseded by the eotitanosuchians and dinocephalian theraspids during the Roadian age. Dimetrodon first appeared in the late Paleozoic Era, and died out following the Permian-Triassic extinction event, 245 million years ago, which immediately preceded the Mesozoic Era. It was more closely related to mammals than to true reptiles such as lizards. When fully grown, Dimetrodon grew up to three meters (9.8 ft) or more and perhaps 150 kg (330 lb), with species varying in weight and size. Dimetrodon is an advanced member of the Sphenacodontid family, and may have evolved from an early Sphenacodon-like form (such as Sphenacodon ferox) during the Late Carboniferous or Early Permian (Gzhelian-Asselian) time.
Iconic and well-known for its immediately recognizable "fin" or "sail" made up of elongate vertebral spines running along its back, up to a meter in length, and in life covered with a layer of skin and blood vessels and serving as a thermoregulatory structure for a heat-exchange mechanism. The sail may have also been used for mating and dominance rituals, and/or for making it look much larger than it was to predators. Despite popular belief, Dimetrodon was not a dinosaur. Rather, it is classified as a pelycosaur, and more closely related to mammals than to reptiles. Dimetrodon was the dominant predator in its environment for nearly twenty-five million years, during which time it evolved into about a dozen species, becoming steadily larger as time progressed.
Dimetrodon fossils have been found in Early to Middle Permian deposits of a localized area in western equatorial Euramerica, now Oklahoma (Wellington Formation) and Texas. A significant find by Jerry MacDonald was the discovery of Dimetrodon footprints in southern New Mexico. Fragmentary remains have also been found in Europe.
Description[]


{kind=link}
Fossil skeleton of Dimetrodon grandis, National Museum of Natural History, Washington, D.C.
Dimetrodon was an apex predator, among the largest of its day. It grew to up to 11.5 feet (3.5 meters) in length, and weighed approximately 550 pounds (250 kg). The name Dimetrodon means 'two-measures of teeth', so named because it had a large skull with two different types of teeth (shearing teeth and sharp canine teeth), unlike reptiles. It walked on four side-sprawling legs and had a large tail. Dimetrodon may have moved in a manner similar to present-day lizards, on four legs that sprawled out to the sides (unlike dinosaurs, whose legs extended under their bodies).
Skull[]
Dimetrodon had a large skull, with a small orbit and high temporal fenestra.[1]
Dentition[]
In the front of it's jaw, Dimetrodon had sharp and chisel-shaped teeth.[2] Behind those teeth are five sharp, triangular cutting teeth, not unlike the sectorial teeth of carnivores like the tiger and lion.[2] Behind those teeth lie simple cones, supposedly useful in holding a struggling victim.[2]
Sail[]
The most distinctive characteristic of Dimetrodon was the spectacular sail on its back (other pelycosaurs such as Edaphosaurus, Ianthasaurus, and Sphenacodon also have this trait). The sail. This sail, which is composed of a row of long spines, each of which grew out of a separate vertebra was supported by neural spine processes, projected upward from its vertebrae. A thin layer of skin was stretched over this elaborate sail. The sail spines were as high as one third the entire length of its body and approached 40 inches high.
Throughout the animal's lifetime, the neural spines continually grew in length. This increase in length is proportional to the animal's increase in body weight, not body length.[1]
Function[]
The sails probably had a display or thermoregulatory function (or both). Dimetrodon and Edaphosaurus lived together, a carnivore and a herbivore, and both developed sails.[3][4] This dynamic would repeat in the Cretaceous with Spinosaurus and Ouranosaurus.The sail, which was dense with blood vessels, was probably used to regulate body temperature; the surface area would allow it to warm up or cool off more efficiently. This adaptation was important because it would give the animal more time to hunt prey. The sail may also have been used in mating rituals and to warn off other predators. The sail was supported by neural spines, each one sprouting from an individual vertebra. Bramwell and Fellgett (1973) calculate that a 200 kg Dimetrodon would heat up from 26° C to 32° C in 205 minutes without a sail and in only 80 minutes with a sail.[3]
History[]
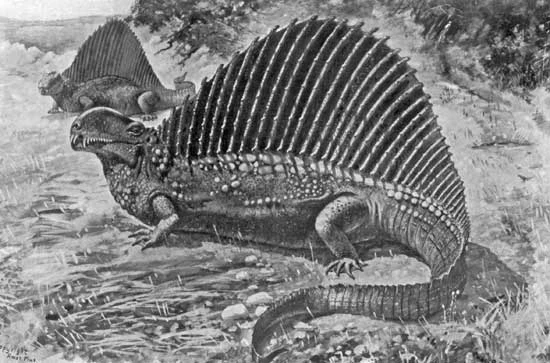

{kind=link}
"Naosaurus" as envisioned by Charles R. Knight. Note the transverse struts on the sail spines indicative of Edaphosaurus.
In 1886, paleontologist Edward Drinker Cope wrote an article in the American Naturalist, "The Long-Spined Theromorpha of the Permian Epoch", in which he described his idea as to the purpose of Dimetrodon's sail:
- "The utility [of the sail] is difficult to imagine. Unless the animal had aquatic habits and swam on its back, the crest or fin must have been in the way of active movements."
It was once thought that the sails may have been sexually dimorphic, and that the female of Dimetrodon was Sphenacodon, but seeing as the two never occurred together (and, if fact, operated in different strata) doesn't give much credence to the idea.
Slight differences in the bony back struts of a similar animal that inhabited the same habitat as Dimetrodon provided paleontologist E.D. Cope with a clue. Whereas Dimetrodon was a large apex predator, "Naosaurus" appeared to be a more peaceful creature. Today it is now known that "Naosaurus" was a chimera, the head of a Dimetrodon attached to the body of the sail-backed herbivore Edaphosaurus, but this was not initially apparent. The bodies of the two animals were similar but since "Naosaurus" lacked a skull it was difficult to be sure what its habits were. In 1908 paleontologist E.C. Case speculated on the habits of "Naosaurus" (attributing to it a more Edaphosaurus-like skull) in Popular Science by stating:
- "In a recent restoration of Naosaurus it has been given the skull of the fiercely carnivorous Dimetrodon, the general similarity of the forms seemed to warrant this, but recent discoveries have made it probable that Naosaurus was not an eater of flesh, but a peaceful, sluggish eater of shell fish and perhaps of vegetation. This animal has perhaps the most wonderful dentition of any known animal the incisor teeth are sharp and chisel-shaped, such as might be useful in cutting strong vegetation; behind these are five sharp triangular cutting teeth, not unlike the sectorial teeth of such flesh eaters as the tiger and lion; behind these simple cones, such as would be useful in holding a struggling victim."
- —E.C. Case
This would make Dimetrodon the dominating carnivore of the time and "Naosaurus" an omnivore, but this only made the puzzle of the sails more complex. A question remained at the time: Why should animals with different habits share the same structure? Multiple explanations had been proposed. The most abused notion was Cope's suggestion that the term "sail" was especially apt; that these animals were using these structures just like a boat sail.[citation needed]
In 1886, Cope wrote a short note on "The Long-Spined Theromorpha of the Permian Epoch" for the American Naturalist. Of the sail on Dimetrodon Cope wrote:
- "The utility [of the sail] is difficult to imagine. Unless the animal had aquatic habits and swam on its back, the crest or fin must have been in the way of active movements."
- —E. D. Cope
However, Cope also noted that the sail of "Naosaurus" had an important difference:
- "There the spines are not quite so elevated as in the [Dimetrodon], but they are more robust, and have transverse processes or branches which resemble the yardarms of a ship's mast. ... The animal must have presented an extraordinary appearance. Perhaps its dorsal armature resembled the branches of shrubs then, as they do now, and served to conceal them in a brushy or wooded region. Or, more probably, the yardarms were connected by membrane with the neural spine or mast, thus serving the animal as a sail with which he navigated the waters of the Permian lakes."
- —E. D. Cope
This concept remained a popular notion[5] but it seems to have been a target of more ridicule than careful consideration. The idea that the sails provided some kind of camouflage was more reasonable, but it seemed strange that a billboard-like structure should serve as a feature that would conceal already-large animals.
Cope's hypotheses had failed, but perhaps there was another explanation. Case wondered if the sails could be the result of a once-useful structure outgrowing its usefulness, the product of some kind of "evolutionary inertia":
- "Such a development seems to have occurred once and again in the history of the world, and the most bizarre types of life owe something of their condition, at least, to this principle. The late Professor Beecher, of Yale, has shown that there is a decided tendency, both in plants and in animals, for a species that is nearing the point of its extinction to develop a spiny or horny habit, covering itself with all sorts of excrescences, seemingly in an unregulated effort to find some condition which will prolong its existence. It is certain that these reptiles, dominant as they were, were rapidly completing their allotted span and as the end approached the spines grew ever heavier and heavier, until it seems plausible to suggest that they became at least a great drain on the animal's powers of nutrition and hastened in no slight degree the end."
- —E.C. Case
At the time, Case was working when non-Darwinian evolutionary mechanism were popular in paleontology, making this language unusual today. It was not strange to think that not only might evolution be directional, but lineages might have a kind of natural timer that determined "their allotted span." In this way lineages were treated like individuals, having a "life" from birth (origin of a new group) to death (extinction). It was as if Dimetrodon and its kind were desperately trying to find some new adaptation to stave off the end.
These sorts of notions were cast out when the modern evolutionary synthesis coalesced in the mid-20th century, but the sails of Dimetrodon, Edaphosaurus, and similarly-endowed creatures from other groups still puzzle paleontologists. Thermoregulation and sexual selection are most often considered to have played a role in the origin of these structures but since it is not possible to directly observe the physiology or behavior of these animals it is difficult to be sure. Ornate sails have independently evolved so many times in the course of vertebrate evolution that questions about their origins and functions are still worthy of consideration.
Paleoecology and Paleobiology[]
Based on associated fossils of other flora and fauna found with Dimetrodon fossils, paleontologists have been able to accurately ascertain the kind of environment that Dimetrodon lived in. The Permian habitat that Dimetrodon lived in had an an arid landscape composed of scrub-like vegetation and often subjected to droughts.
Infection and disease[]
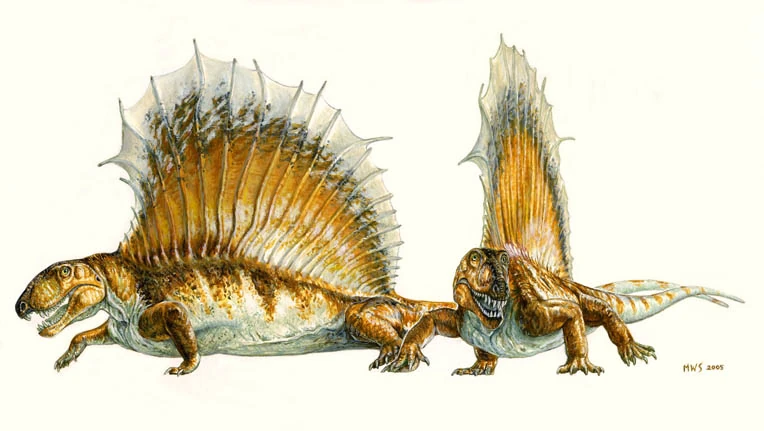

{kind=link}
Artist's impression of a D. gigashomogenes with a pathological sail.
The sail of Dimetrodon was just as likely to be subject to injury and disease just like any other bones. A particular specimen of Dimetrodon gigashomogenes, described by Stuart Sumida and Elizabeth Rega and restored by Skrepnick, had suffered some injury earlier in life that caused some of the spines supporting the sail to break. When the bones healed they did not retain their "normal" position. Instead they twisted and bent forward. The pathologies occur along several anterior dorsals, proximally on the spines (close to the centrum )and rehealing/exostosis is present on the bone surface. However, conversely and in addition to these injuries, the distal ends of many of the spines throughout the entire sail series are severely bent and twisted. Furthermore, because of the arrangement of the spines relative to each other, as the specimen was originally discovered in situ, this peculiar deformation in spine morphology is not an artifact of postdepositional/geological forces at work, but rather seems to have been "natural" to the animal in life.[6]
Whereas many specimens of Dimetrodon have relatively conservative, orderly sail spine arrangements (although some seem to show some a bit of warp that might be ordinarily discounted to crushing/bending postmortem) this individual shows that there may have been an interesting variation of "displays" in these sails, not previously considered.[6]
Another specimen of Dimetrodon discovered in the Texas Red beds exhibited an incompletely healed fracture of the left radius. The bone exhibited a great deal of osteosclerosis and some intermediary callus. This is the oldest known case of a fracture represented in the fossil record. The fracture was described as a "simple one", with the bone being broken at right angles and healing began shortly afterwards. The bone also showed no signs of medullary activity, rendering x-rays ineffective in this case.[7]
Relationship with modern mammals[]


{kind=link}
Dimetrodon incisivus attacking a Varanosaurus acutirostris.
As a synapsid, Dimetrodon was distantly related to modern mammals. Synapsids were the first tetrapods to evolve differentiated (or heterodont) teeth. Whereas reptiles hardly chew their food, simply gulping it down, synapsids like Dimetrodon developed teeth to help shear meat into smaller pieces for easier ingestion. These 'two-measure teeth' eventually gave rise to the various kinds of teeth present in modern mammals.
Dimetrodon's protruding sail also allowed the synapsid to partially regulate its body temperature, which marked may have marked a transition into endothermic mammals.
Gallery[]
")
")
")
See also[]
- Edaphosaurus — a herbivore pelycosaur with a sail on its back.
- Haptodus — a primitive sphenacodont.
- Sphenacodon — a pelycosaur closely related to Dimetrodon.
- Platyhystrix — an unrelated animal with a sail on its back.
- Evolution of mammals
References[]
- ^ a b Benton, Michael J. Vertebrate Palaeontology. Blackwell Publishers. 2nd edition (June 2000) pp. 113-115 ISBN-10: 0632056142
- ^ a b c Palmer, D., ed (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. p. 187. ISBN 1-84028-152-9.
- ^ a b Bramwell, C. D. and Fellgett, P. P., 1973, Thermal regulation in sail lizards. Nature, v. 242, p. 203-205.
- ^ Both came from Permian strata in Texas.
- ^ It was featured in a 1897 article on "Strange Creatures of the Past" in The Century, among some other ideas
- ^ a b A Dimetrodon with a funky sail Science Blogs article by Laelaps.
- ^ Brothwell, Don 1967. Diseases in Antiquity: A Survey of the Diseases, Injuries, and Surgery of Early Populations (Hardcover). Charles C Thomas Pub Ltd. ISBN: 0398002339