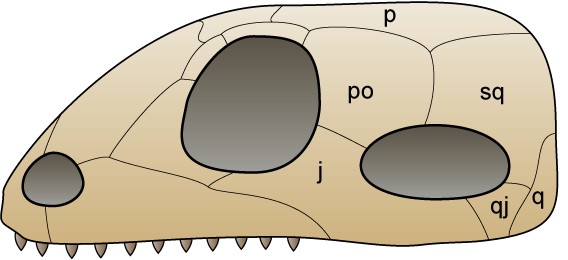
Synapsids ('fused arch'), also known as theropsids ('beast eye'), are a clade of animals that includes mammals and everything closer to mammals than to other living amniotes.[1] The non-mammalian members were traditionally described as mammal-like reptiles,[2][3] and are sometimes referred to as "proto-mammals" or "stem-mammals". Synapsids are one of the two major groups of amniote, the other being the sauropsids (or reptiles in the proper sense). They are distinguished from the latter by a single opening (temporal fenestra) in their skull behind each eye, which developed in the ancestral synapsid about 324 million years ago (mya) during the lateCarboniferousPeriod.
Synapsids were the dominant terrestrial animals in the middle to late Permian period. As with almost all life forms then extant, their numbers and variety were severely reduced by the Permian extinction. Some species survived into the Triassic period, but archosaurs quickly became the dominant animals and few of the non-mammalian synapsids outlasted the Triassic, although survivors persisted into the Cretaceous. However, they included the prehistoric ancestors of mammals and in this sense, synapsids are still very much a living class of vertebrates.
Synapsids were originally defined at the turn of the 20th century, as one of the four main subclasses of reptiles, on the basis of their distinctive temporal openings. These openings in the cheek bones allowed attachment of larger jaw muscles, hence a more efficient bite. Synapsids were considered to be the reptilian lineage that led to mammals via gradually evolved, increasingly mammalian features, hence the name "mammal-like reptiles" which became a broad, traditional description for all non-mammalian synapsids.[2][3]
The traditional classification continued through to the late 1980s (e.g. Carroll 1988). In the 1990s this approach was replaced by a cladistic one, according to which the only valid groups are those that include common ancestors and all of their descendants: these are known as monophyletic groups, or clades. Because mammals are directly descended from (other) synapsids, mammals are included under Clade Synapsida. However, formal classification has not kept pace with cladistics, so mammals are still often treated as a separate class alongside a paraphyletically-defined Class Synapsida. At the same time, the term "reptiles", which traditionally denoted all cold-blooded amniotes, is now re-defined to include only the sauropsids (a class that unites the anapsids and the diapsids to the exclusion of the synapsids, because these first two groups are more closely related to each other than to the third one). Hence the term "mammal-like reptiles" for the synapsids is considered obsolete under this terminology.
The non-mammalian synapsids are traditionally divided into a primitive and an advanced group, known respectively as 'pelycosaurs' and therapsids. 'Pelycosaurs' are an artificial, paraphyletic grouping of six families which are united only in that they are primitive in relation to therapsids, and do not constitute a clade.[4] They are currently divided between two primary synapsid clades, the Caseasauria and the Eupelycosauria, the latter of which also includes all the more advanced synapsids and therefore the mammals. That is to say, therapsids make up a well-defined clade within the eupelycosaurs, as long as mammals are included in the therapsids.
Synapsids evolved a temporal fenestra behind each eye orbit on the lateral surface of the skull. It may have evolved to provide new attachment sites for jaw muscles.
Some synapsids (including mammals) also have a warm-blooded metabolism, even though early synapsids, such as pelycosaurs, are believed to have been cold-blooded.
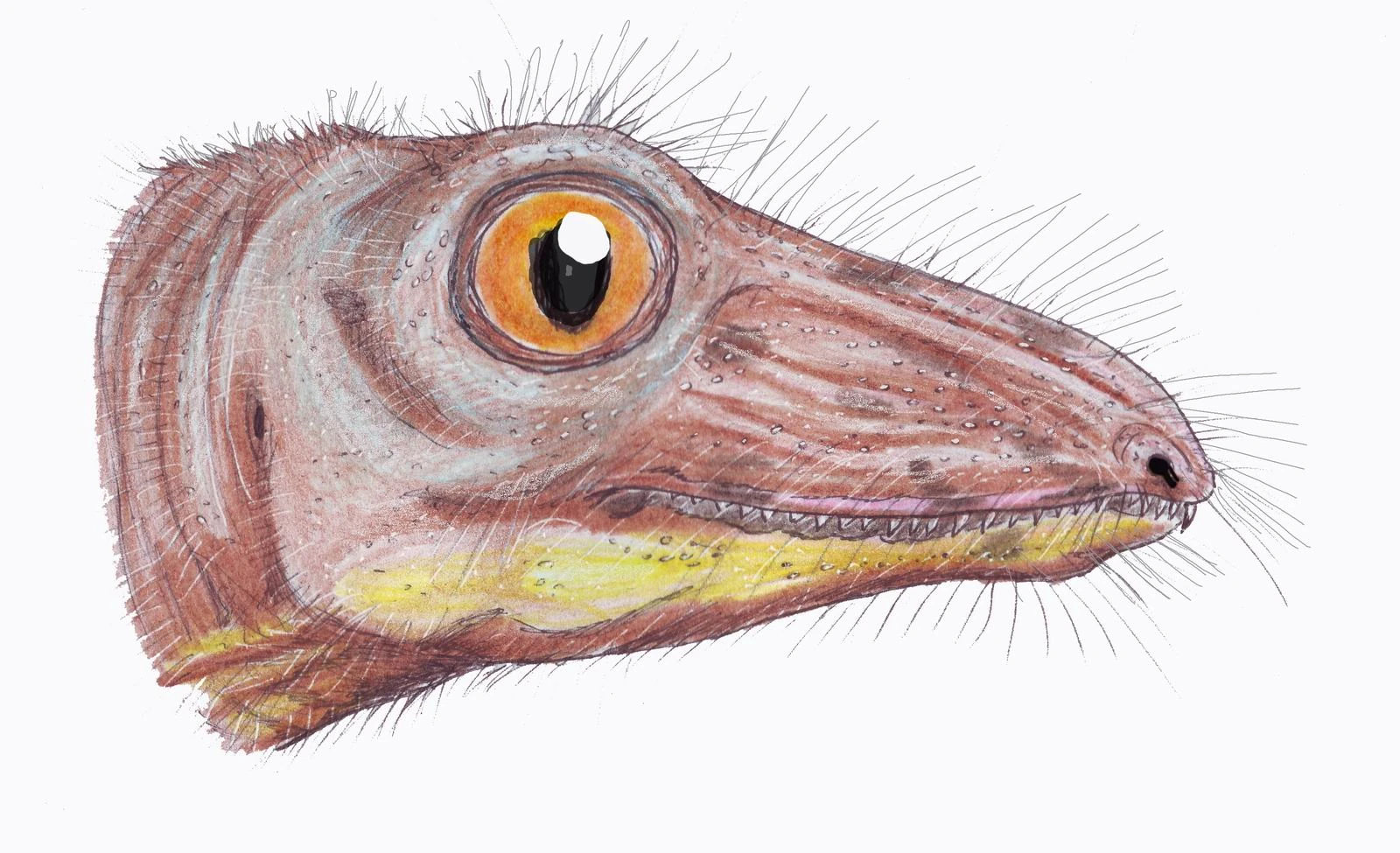
Like mammals, the nonmammalian synapsids possessed glandularskin that lacked scales, though at least the pelycosaurs retained the "scales" of more primitive tetrapods on their undersides. These scales differed in structure from reptilian scales, an epidermal feature like mammalian hair or avian feathers.[5] It is currently unknown at what stage they acquired mammalian characteristics like body hair and mammary glands, as the fossils only rarely provide direct evidence for soft tissues. Much, however, can be inferred from differences in skeletal structure. Thus the more primitive synapsids can be better visualized as being "naked lizards", both furless and scaleless, as their overall aspect was more like a modern lizard than a modern mammal, and the distinguishing features are relatively fine ones of internal structure. On the other hand, the presence of a secondary palate, erect posture and other indicators of high metabolic rate among the advanced cynodonts strongly suggests that many mammalian features, including an effective insulating layer of body hair, had evolved by this stage. This is now confirmed by impressions of fur in rocks directly underlying some fossil therapsids.[6]
Synapsids are the first tetrapods to have differentiated teeth. These include the canines, molars, and incisors. Early synapsids had multiple jaw bones. As they evolved, these jaw bones were reduced in size and gradually moved into the ear, forming the middle ear bones.
Most paleontologists hold fossilized jaw remains to be the distinguishing feature used to classify synapsids and reptiles. The jaw transition is a good classification tool as most other fossilized features that make a chronological progression from a reptile-like to a mammalian condition follow the progression of the jaw transition. The dentary, or lower jaw, consists of a single bone in mammals, where the lower jaw of modern and prehistoric reptiles consists of a conglomeration of smaller bones.
Mammalian jaw structures are also set apart by the dentary-squamosal jaw joint. In this form of jaw joint, the dentary forms a connection with a depression in the squamosal known as the glenoid cavity. In contrast, all other jawed vertebrates, including reptiles and nonmammalian synapsids, possess a jaw joint in which one of the smaller bones of the lower jaw, the articular, makes a connection with a bone of the skull called the quadrate to form the articular-quadrate jaw joint. In forms transitional to mammals, the jaw joint is composed of a large, lower jaw bone (similar to the dentary found in mammals) that does not connect to the squamosal but connects to the quadrate with a receding articular bone.
Over time, as synapsids became more mammalian and less 'reptilian', they began to develop a secondary palate, separating the mouth and nasal cavity. In early synapsids, a secondary palate began to form on the sides of the maxilla, still leaving the mouth and nostril connected.
Eventually, the two sides of the palate began to curve together, forming a U-shape instead of a C-shape. The palate also began to extend back toward the throat, securing the entire mouth and creating a full palatine bone. The maxilla is also closed completely. In fossils of one of the first eutheriodonts, the beginnings of a palate are clearly visible. The later Thrinaxodon has a full and completely closed palate, forming a clear progression.[7]
Archaeothyris and Clepsydrops are the earliest known synapsids.[8] They lived in the Pennsylvanian subperiod of the Carboniferous Period and belonged to the series of primitive synapsids which are conventionally grouped as 'pelycosaurs'. The 'pelycosaurs' were the first successful group of amniotes, spreading and diversifying until they became the dominant large terrestrial animals in the latest Carboniferous and Early Permian Periods. They were sprawling, bulky, cold-blooded and had small brains. They were the largest land animals of their time, ranging up to 3 m (10 ft) in length. Many, like Dimetrodon, had large sails that may have helped raise their body temperature. A few relict groups lasted into the later Permian, but most of the 'pelycosaurs' became extinct before the end of Permian.
The therapsids, a more advanced group of synapsids, appeared during the first half of the Permian and went on to become the dominant large terrestrial animals during the latter half. They have dominated the world twice: once in the Permian and once in the Cenozoic, the current era. They were by far the most diverse and abundant animals of the Middle and Late Permian and included herbivores and carnivores, ranging from small animals the size of a rat (e.g: Robertia), to large bulky herbivores a ton or more in weight (e.g: Moschops). After flourishing for many millions of years, these successful animals were all but wiped out by the Permian-Triassic mass extinction about 250 Mya, the largest extinction in Earth's history, which may have been related to the Siberian Traps volcanic event.
Nikkasaurus - an enigmatic synapsid from the Middle Permian of Russia.
Only a few therapsids (and some relict 'pelycosaur' taxa) survived the Permian extinction and went on to be successful in the new early Triassic landscape; they include Lystrosaurus and Cynognathus, the latter of which appeared later in the early Triassic. Now, however, they were accompanied by the early archosaurs (formerly known as thecodonts; this term is not used in modern classifications). Some of these, like Euparkeria, were small and lightly built, while others, like Erythrosuchus, were as big as or bigger than the largest therapsids.
Triassic therapsids included three groups. Specialised, beaked herbivores known as dicynodonts (such as Lystrosaurus and its descendants, the Kannemeyeriidae), contained some members which reached large size (up to a tonne or more). The increasingly mammal-like carnivorous, herbivorous, and insectivorous cynodonts included the eucynodonts from the Olenekian age, an early representative of which was Cynognathus. Finally, there were the therocephalians, which only lasted into the early part of the Triassic.
Unlike the dicynodonts, which remained large, the cynodonts became progressively smaller and more mammal-like as the Triassic progressed. From the most advanced and tiny cynodonts, which were only the size of a shrew, came the first mammal precursors, during the Carnian age of the Late Triassic, about 220 Mya.
During the evolutionary succession from early therapsid to cynodont to eucynodont to mammal, the main lower jaw bone, the dentary, replaced the adjacent bones. Thus, the lower jaw gradually became just one large bone, with several of the smaller jaw bones migrating into the inner ear and allowing sophisticated hearing.
Whether through climate change, vegetation change, ecological competition, or a combination of factors, most of the remaining large cynodonts (belonging to the Traversodontidae) and dicynodonts (of the family Kannemeyeriidae) had disappeared by the Norian age, even before the Triassic-Jurassic extinction event that killed off most of the large non-dinosaurian archosaurs. The remaining Mesozoic synapsids were small, ranging from the size of a shrew to the badger-like mammal Repenomamus.
During the Jurassic and Cretaceous, the remaining non-mammalian cynodonts were small, such as Tritylodon. No cynodont grew larger than a cat. Most Jurassic and Cretaceous cynodonts were herbivorous, though some were carnivorous. The family Trithelodontidae first appeared near the end of the Triassic. They were carnivorous and persisted well into the Middle Jurassic. The other, Tritylodontidae, first appeared at the same time as the trithelodonts, but they were herbivorous. This group became extinct at the end of the Early Cretaceous epoch. Dicynodonts are thought to have become extinct near the end of the Triassic period, but there is evidence that this group survived. New fossil finds have been found in the Cretaceous rocks of Gondwana.
Today, there are 5,400 species of living synapsids known as the mammals, including both aquatic (whales) and flying (bats) species, and the largest animal ever known to have existed (the blue whale). Humans are synapsids as well. Uniquely among the synapsids, however, most mammals are viviparous and give birth to live young rather than laying eggs, the exception being the monotremes.
Synapsids' evolution into mammals is believed to have been triggered by moving to a nocturnalniche. Proto-mammals with higher metabolic rates were able to keep their bodies warm at night, and were more likely to survive. This meant consuming food (generally thought to be insects) in much greater quantity. To facilitate rapid digestion, proto-mammals evolved mastication (chewing) and specialized teeth that aided chewing. Limbs also evolved to move under the body instead of to the side, allowing proto-mammals to breathe more efficiently during locomotion[9] and also to be able to change direction more quickly in order to catch small prey at a faster rate. This helped make it possible to support their higher metabolic demands. It is believed that, rather than out-running predators, proto-mammals adapted the strategy of outmaneuvering predators using their improved locomotor capabilities.[6]
Bramble, D. M.; Jenkins, F. A. (1993). "Mammalian locomotor-respiratory integration: Implications for diaphragmatic and pulmonary design". Science262 (5131): 235–240. doi:10.1126/science.8211141.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


{kind=link}